
 Verlauf
Verlauf

 Alle Kapitel anzeigen
Alle Kapitel anzeigen voriges Kapitel
voriges Kapitel
Netzhaut, Rezeptorzellen (Stäbchen und Zapfen)
Netzhaut
Die Netzhaut besteht aus mehreren Zellschichten. Die hinterste Schicht bilden die Rezeptorzellen (Stäbchen, Zapfen), die das auftreffende Licht in neuronale Signale umwandeln. Zwischen den Rezeptorzellen und den Ganglienzellen liegt ein Netzwerk von drei Neuronentypen (= Nervenzellen), den Horizontal-, Bipolar- und Amakrinzellen. Diese verbinden die Rezeptorzellen untereinander, sowie die Rezeptorzellen über diverse Verschaltungen mit den Ganglienzellen. In diesem Netzwerk findet durch komplexe Interaktionen eine Vorverarbeitung der Erregungswerte der einzelnen Rezeptoren statt. Die vorderste Schicht bilden die Ganglienzellen, die über die Bipolarzellen direkt sowie über das ganze Netzwerk indirekt mit den Rezeptorzellen in Verbindung stehen.
D.h. das einfallende Licht durchdringt zuerst die Schicht der Ganglienzellen und des Neuronennetzwerks bevor es auf die Rezeptorzellen trifft.
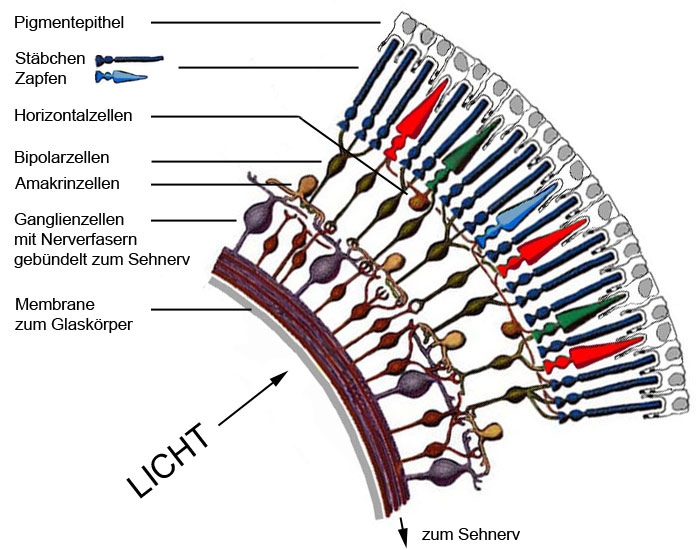
Abb.: Aufbau der Retina (Netzhaut).
Die Nervenfasern (Axione) der Ganglienzellen werden im Sehnerv gebündelt und sind die Verbindung zum Gehirn. Die Anzahl der Ganglienzellen in der Netzhaut beträgt ca. 1,5 Millionen. Das entspricht auch der Anzahl der Nervenfasern im Sehnerv und der Anzahl der Bildpunkte, die das Gehirn erreichen. Eine vergleichsweise geringe Anzahl, wenn man bedenkt dass bei den Digitalfotokameras eine Auflösung von 6 Millionen Pixel bereits Standard ist. Allerdings ist das Signal im Sehnerv bereits vorverarbeitet und das Signal jeder Nervenfaser enthält mehr Information als ein Pixel eines Digitalkamerabildes.
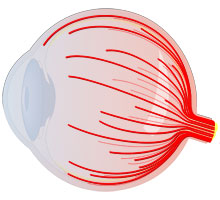
Abb.: Die Nervenfasern der Ganglienzellen überziehen die gesamte Netzhaut und werden im Sehnerv gebündelt.
Rezeptorzellen
Bei den Rezeptorzellen unterscheidet man zwischen zwei Typen, den Stäbchen und Zapfen, die wiederum in S-Zapfen, M-Zapfen und L-Zapfen unterteilt sind. Beide Typen haben im Wesentlichen dieselbe Struktur und enthalten ca. 10 Millionen lichtempfindliche Photopigmentmoleküle, die permanent neu gebildet werden. Jedes Photopigmentmolekül ist zusammengesetzt aus dem lichtempfindlichen Retinalmolekül (ähnlich dem Vitamin-A) und dem Protein Opsin. Werden die Retinalmoleküle von Lichtphotonen der richtigen Wellenlänge getroffen, erfolgt in der Netzhaut über einen mehrstufigen komplexen Vorgang die Umwandlung in einen Nervenimpuls. Dieser Umwandlungsprozess dauert ca. 50 Mikrosekunden (50 millionstel Sekunden).
Das Photopigmentmolekül der Stäbchen heißt Rhodopsin. Die Photopigmentmoleküle der Zapfen unterscheiden sich vom Rhodopsin und untereinander durch leicht veränderte Aminosäuresequenzen im Opsinteil. Dadurch ergibt sich die unterschiedliche Wellenlängenempfindlichkeit.
Stäbchen
Die Stäbchen sind "farbenblind" und können nur Lichtintensitäten verarbeiten. Mit ihnen ist nur eine Hell-Dunkelwahrnehmung (scoptopisches Sehen) möglich. Sie arbeiten bereits bei sehr geringen Beleuchtungsstärken (Dämmerungssehen). Die Gesamtanzahl der Stäbchen beträgt ca. 120 Millionen.
Zapfen
Die Zapfen ermöglichen Farbsehen und Hell-Dunkelwahrnehmung (photopisches Sehen). Weil es drei verschiedene Zapfentypen gibt, die unterschiedliche Empfindlichkeitsbereiche und -maxima innerhalb des sichtbaren Spektrums aufweisen, können aus der Kombination der drei Erregungsinformationen nicht nur wie bei den Stäbchen Lichtintensitäten unterschieden werden, sondern auch verschiedene Wellenlängen (= Farben). Die einzelnen Zapfentypen sind isoliert betrachtet wie die Stäbchen "farbenblind".
Die Zapfen benötigen eine relativ hohe Lichtstärke und sind bei Tageslicht wirksam. Sie brauchen etwa 30-fach höhere Lichtintensität als die Stäbchen. Die Gesamtanzahl der Zapfen beträgt ca. 6 Millionen.

Stäbchen (groß) und Zapfen (klein und gelb eingefärbt) in einer elektronenmikroskopischen Aufnahme der Retina eines Schwanzlurchs. Das Pigmentepithel ist abpräpariert.
(elektronenmikroskopische Aufnahme entnommen aus http://www.sinnesphysiologie.de/hvsinne/auge/rodcone.htm)
Empfindlichkeitsbereich der Stäbchen und Zapfen
Stäbchen werden optimal von blau-grünem Licht (mit einer Wellenlänge von ca. 500 nm) angeregt und vermitteln die Wahrnehmung von Grautönen.
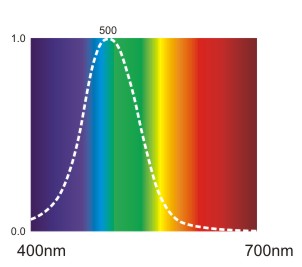
Abb.: Normalisierte Sensitivitätskurve der Stäbchen.
Die Kurve ist eine normalisierte Darstellung, die für jede Wellenlänge die Sensitivität der Stäbchen angibt. Dabei wird die maximale Sensitivität, die bei 500 nm liegt, mit 100% oder 1.0 angenommen und relativ dazu werden die Erregungswerte bei anderen Wellenlängen in Prozent ausgedrückt. Für die Erstellung der Kurve ist es natürlich wichtig, dass für jede Wellenlänge Licht derselben Intensität auf die Stäbchen trifft. Licht, das nur aus einer Wellenlänge "aufgebaut" ist, nennt man monochromatisches Licht.
Zapfen dagegen teilen sich das Spektrum in drei Bereiche auf: Das Absorptionsmaximum der S-Zapfen (short wavelength) oder Blauzapfen liegt bei ca. 420 nm, das der M-Zapfen (medium wavelenght) oder Grünzapfen bei ca. 540 nm und das der L-Zapfen (long wavelength) oder Rotzapfen bei ca. 560 nm (wobei eigentlich das Empfindungsmaximum der Rotzapfen im gelben Bereich liegt). [Die nm-Werte schwanken leicht in der Literatur.]
Die Information, welche Zapfenart wie stark angeregt wird, ist die Ausgangsbasis für einen komplexen Informationsverarbeitungsprozess in der Retina und im Gehirn, der in einer Farbempfindung mündet.
In der nachstehenden Abbildung erkennt man, dass die S-Zapfen nur auf Licht von violett bis gelbgrün, die M- und L-Zapfen jedoch auf Licht aus dem gesamten Spektrum reagieren. Die M- und L-Zapfen haben sehr ähnliche Empfindungskurven, da sich ihre Photopigmentmoleküle kaum unterscheiden. Die L-Zapfen werden im Rotbereich stärker angeregt .
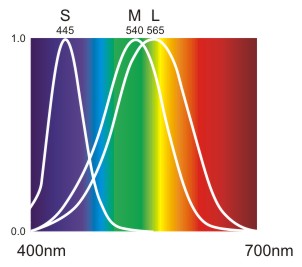
Abb.: Normalisierte Absorptionskurven (Sensitivitätskurven) der drei Zapfentypen.
Diese Kurven liefern die relative Erregung der Rezeptoren bei einfallendem Licht einer bestimmten Wellenlänge. D.h. man isoliert die Rezeptoren, setzt sie dem Licht einer bestimmten Wellenlänge aus und misst die Erregung. So erhält man die Erregungswerte für jede Wellenlänge, wobei wichtig ist, dass für jede Wellenlänge dieselbe Lichtintensität verwendet wurde. Dem höchsten Erregungswert wird 100% bzw. 1.0 (= Empfindungsmaximum) und allen anderen Wellenlängen werden die entsprechenden Prozentwerte zugeordnet.
Zu beachten ist, dass diese Kurven keinen Vergleich der absoluten Empfindlichkeit der drei Zapfentypen bieten. Für einen Vergleich ist es wichtig, dass es gemessen an der Gesamtzahl der Zapfen 63% L-Zapfen, 31% M-Zapfen und nur 6% S-Zapfen gibt. Daraus ergibt sich nachstehendes vergleichendes Kurvenbild.
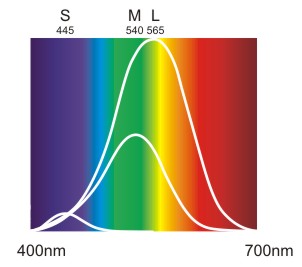
Abb.: Normalisierte Absorptionskurven der drei Zapfentypen gewichtet nach der prozentuellen Verteilung.
Es gibt ungefähr doppelt soviele L-Zapfen wie M-Zapfen und fünf mal mehr M-Zapfen als S-Zapfen. Daher ist das Maximum der L-Kurve doppelt so hoch wie bei der M-Kurve und deren Maximum fünf mal höher als bei der S-Kurve.
Diese Kurven sagen jedoch nichts darüber aus, wie die Informationen von den Zapfen weiterverarbeitet werden. Denn tatsächlich kommt den Informationen von den S-Zapfen wesentlich mehr Bedeutung zu als es in dieser Abbildung zu vermuten wäre.
Verteilung von Stäbchen und Zapfen auf der Netzhaut
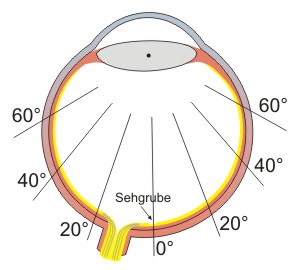
Abb.: In Texten liest man von Gradangaben, wenn Bereiche der Netzhaut beschrieben werden. 0° steht dabei für die Sehachse von der Linse zum Zentrum der Sehgrube.
Die Stäbchen und einzelnen Zapfentypen sind auf der Netzhaut sehr unterschiedlich verteilt.
Die Sehgrube (Fovea, ca. 1 mm) enthält nur schlanke Zapfen in einer sehr dichten regelmäßigen hexagonalen Anordnung. Im Zentrum der Sehgrube (Foveola) gibt es nur M- und L-Zapfen (bis zu 250.000 pro mm²), die zum Sehgrubenrand auf 50.000 pro mm² und weiter zum Rand der Netzhaut auf 5.000 pro mm² abnehmen. Die S-Zapfen, die im Zentrum der Sehgrube vollkommen fehlen, haben ihre größte Dichte am Sehgrubenrand (bei 1°) mit 2.000 pro mm² und nehmen zum Rand der Netzhaut auf 500 pro mm² ab.
Jeder Rezeptorzelle in der Sehgrube ist eine Ganglienzelle zugeordnet. Dadurch wird eine hohe Auflösung erreicht. Die Sehgrube ist somit der Bereich des schärfsten Sehens auf der Netzhaut. Ein Gegenstand, welcher vom Betrachter fixiert wird, wird immer so abgebildet, dass dessen Abbild genau in der Sehgrube, dem Bereich des schärfsten Sehens liegt. Ungefähr die Hälfte aller Zapfen ist im Bereich der Sehgrube angeordnet.
Da durch die geringe Dichte der peripheren Zapfen und der Krümmung der Netzhaut kein scharfes Bild möglich ist, dienen die Zapfen in den Außenbereichen der Netzhaut für die Wahrnehmung von Bewegung, Helligkeit und Farbkontrast. Wir können am Rand unseres Gesichtsfeldes (im Augenwinkel) zwar Bewegungen, aber keine genauen Formen wahrnehmen.
Die Stäbchen, die in der Sehgrube nicht vorhanden sind, haben ihre größte Dichte von ca. 150.000 pro mm² in einem Ring um die Sehgrube (bei 17°) und nehmen zum Netzhautrand auf 80.000 pro mm° ab. Daher darf man im Dämmerlicht einen Gegenstand nicht fixieren (in der Sehgrube sind keine Stäbchen), sondern muss den Blick neben dem Gegenstand ausrichten, damit dieser auf dem Stäbchenring abgebildet wird.
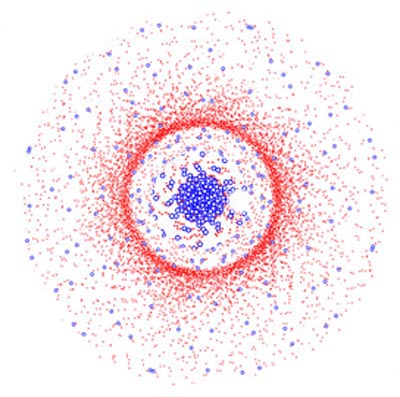
Abb.: Schematische Darstellung der Verteilung von Zapfen (blaue Kreise) und Stäbchen (rote Punkte) auf der Netzhaut. Man erkennt deutlich die Zapfendichte in der Sehgrube und den Stäbchenring.
Rechts von der Sehgrube ist der weiße Kreis des blinden Flecks zu erkennen, wo keine Rezeptorzellen liegen können.